
人類にウイルスが、一体いつ頃から感染するようになったかを知るために使われる一般的なやり方があります。ウイルスは宿主細胞に感染して増殖するために必要な全ての遺伝子情報を担った塩基配列からなる核酸(DNAあるいはRNA)が主体となっていますが、塩基が常に入れ替わって変異しています。1つの位置(サイトといいます)あたり、1年間に塩基が何回(1万分の1個とか10万分の1個というような小さな数字になります)入れ替わるのかがわかれば、変異の速度が計算できます。勿論、その速度が一定で何千年も、何万年も変わらないことが前提になります。たとえば3,000個の塩基からできているウイルスで、日本で感染している患者さんから得られたウイルスとアメリカの患者さんのウイルスとの塩基配列が3箇所で違っていたとします。変異の確率が塩基1個あたり年間1万分の1としますと、3,000個のうち3個の塩基が変化するのに何年(y年)かかるかが計算できます:
3,000 × 1/10,000 × y = 3
の計算式からy = 10、すなわち10年という答えが得られます。同じウイルスが10年間でそれだけ違うように進化したので「アメリカから日本へ、あるいは日本からアメリカにそのウイルスが到来したのは10年以前だった」と推論できます。
ウイルスの変異速度を知るためには、同一の宿主で感染した時点とそれから何年も、あるいは何10年もたってから得られた2個のウイルスで塩基配列を調べて、塩基が何個違うかを調べる必要があります。したがって、持続感染するウイルスであることが条件となります。すなわち肝炎ウイルスでは、血液を介して感染するB型肝炎ウイルスとC型肝炎ウイルスだけが対象となり、糞口感染の経路で一過性に感染し急性肝炎を起こすが、慢性肝炎は起こさないA型肝炎ウイルスとE型肝炎ウイルスでは、かなり速いことは想像できるのですけれども、進化の基準になる変異速度を計算する手段がありません。
C型肝炎ウイルスが人類に感染するようになったのは、比較的最近のことでせいぜい200年以前(19世紀)であろうと考えられています。主に母児感染によって先祖から代々引き継がれたB型肝炎ウイルスは、もっともっと古く有史前からだろうと推定できるのですが、そもそも最初にいつ、そして何処でどの動物から人類がB型肝炎ウイルスに感染したかに関しては、専門家の間でも意見が分かれています。限られた手がかりを元にして、B型肝炎ウイルスの起源を探る作業には大きな夢と深いロマンがあります。これからご一緒に、全世界人口の6%に相当する3億5千万人もの人々が持続感染しているB型肝炎ウイルスのルーツを求め、無限の過去にさかのぼってタイム・トラベルにでかけましょう。
遺伝情報
 地球上に生存するありとあらゆる生物は、たった4種類の塩基の組み合わせからできている遺伝子に記入された設計図にしたがって生活し、それを子孫に遺しています(図1)。塩基はプリンに属するアデニン(A)とグアニン(G)およびピリミジンの構造をもつチミン(T)とシトシン(C)とに分かれます。人間に代表される動物の遺伝子は二重らせんの構造をしています。らせん階段の各ステップがA・TとG・Cの結合からできていて、この組み合わせは発見者の名前にちなんでワトソン・クリック結合と云われています。この結合のために、プリン分子(AとG)の総数とピリミジン分子(TとC)の総数は、二重らせん構造の遺伝子をもつ動物では、すべて同じになります。
地球上に生存するありとあらゆる生物は、たった4種類の塩基の組み合わせからできている遺伝子に記入された設計図にしたがって生活し、それを子孫に遺しています(図1)。塩基はプリンに属するアデニン(A)とグアニン(G)およびピリミジンの構造をもつチミン(T)とシトシン(C)とに分かれます。人間に代表される動物の遺伝子は二重らせんの構造をしています。らせん階段の各ステップがA・TとG・Cの結合からできていて、この組み合わせは発見者の名前にちなんでワトソン・クリック結合と云われています。この結合のために、プリン分子(AとG)の総数とピリミジン分子(TとC)の総数は、二重らせん構造の遺伝子をもつ動物では、すべて同じになります。
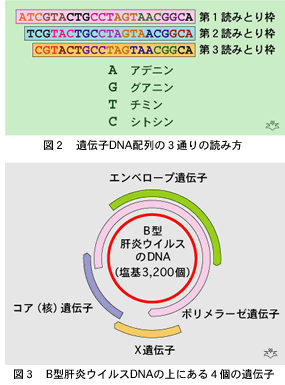 4種類の塩基が連なってできている遺伝子は、生命の維持に必要な体の成分であるタンパクを合成する情報を担っています。タンパクは約20種類のアミノ酸がいろいろな組み合わせで連なってできたアミノ鎖が、立体構造をもち特有の形となったものです。アミノ酸一つずつが3個の塩基の並び方(順列)で決まります。ですから、一つの塩基配列にも、読み始めを一つずつずらすことによって、3種類の違ったタンパクの設計図を書き込むことができます(図2)。沢山の大きな遺伝子をもった高等生物では、使っていない遺伝子も沢山ありますのでその必要がありませんが、限られた数の遺伝子しか持っていない小さな病原体であるウイルスは、同一の塩基配列の上で読みとり枠をずらすことによって複数のタンパクを宿主細胞に合成させることができます。B型肝炎ウイルスがその典型例です。塩基が約3,200個しかない最小のDNAウイルスで、ウイルスの「衣(エンベロープ)」をつくるための遺伝子、核酸を含むウイルスの「コア(核)」をつくる遺伝子と、核酸配列全体の複製(ウイルスの増殖)に必要な酵素(DNAポリメラーゼと逆転写酵素)を合成するための遺伝子を持っています。そのほかに働きがまだわかっていない“Xタンパク”を作るためのX遺伝子もあり合計4個の遺伝子が一本の環になった3,200個の塩基配列の上に乗っています(図3)。これだけ沢山の遺伝子情報を極端に短い塩基配列で伝えるために、一カ所で二つの遺伝子を担っている場所が、B型肝炎ウイルスでは全長の半分位もあります。
4種類の塩基が連なってできている遺伝子は、生命の維持に必要な体の成分であるタンパクを合成する情報を担っています。タンパクは約20種類のアミノ酸がいろいろな組み合わせで連なってできたアミノ鎖が、立体構造をもち特有の形となったものです。アミノ酸一つずつが3個の塩基の並び方(順列)で決まります。ですから、一つの塩基配列にも、読み始めを一つずつずらすことによって、3種類の違ったタンパクの設計図を書き込むことができます(図2)。沢山の大きな遺伝子をもった高等生物では、使っていない遺伝子も沢山ありますのでその必要がありませんが、限られた数の遺伝子しか持っていない小さな病原体であるウイルスは、同一の塩基配列の上で読みとり枠をずらすことによって複数のタンパクを宿主細胞に合成させることができます。B型肝炎ウイルスがその典型例です。塩基が約3,200個しかない最小のDNAウイルスで、ウイルスの「衣(エンベロープ)」をつくるための遺伝子、核酸を含むウイルスの「コア(核)」をつくる遺伝子と、核酸配列全体の複製(ウイルスの増殖)に必要な酵素(DNAポリメラーゼと逆転写酵素)を合成するための遺伝子を持っています。そのほかに働きがまだわかっていない“Xタンパク”を作るためのX遺伝子もあり合計4個の遺伝子が一本の環になった3,200個の塩基配列の上に乗っています(図3)。これだけ沢山の遺伝子情報を極端に短い塩基配列で伝えるために、一カ所で二つの遺伝子を担っている場所が、B型肝炎ウイルスでは全長の半分位もあります。
変異速度を正確に反映する塩基配列の部位
B型肝炎ウイルスに持続感染していて、何10年も以前の血清が保存されているキャリアで新旧2個のウイルス核酸配列を比べることによって、塩基一カ所あたりの年間変異率を計算することができます。正確な変異率を知るために、いくつか考えておかなければならないことがあります。当然のことですが、2個の遺伝子を担っている場所では、1個の遺伝子しか乗っていない部位と比べて、変異がずっと起きにくいのです。遺伝子が2個乗っている場所で、ある位置の塩基が1個替わったとします。その変異がたとえ2個の遺伝子の一方が指令して作るタンパクを構成するアミノ酸に影響しなかったとしても、他方のタンパクのアミノ酸配列を変えてしまうからです。このためにB型肝炎ウイルスでは、正確な変異率を計算できる場所は限られてしまいます。元々総塩基数が少ないので、変異率を知ることがますます困難になります。
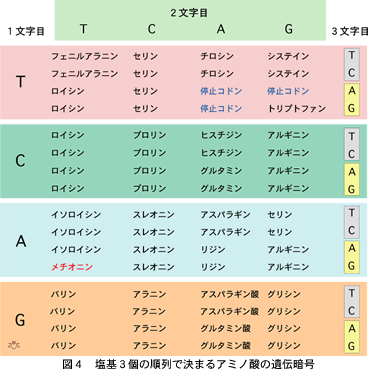 このほかにも、考えておかなければいけないことがあります。3個の塩基の順列が20個のアミノ酸を規定していて、これを“コドン”といっています。一文字目、二文字目と三文字目にそれぞれT、C、AとGの4通りがありますから、4を3回掛けた回数、すなわち64個の組み合わせがあります(図4)。アミノ酸の総数は20個しかありませんから、殆どのアミノ酸に複数のコドンが対応し、中には同じアミノ酸の合成を指令するコドンが6種類もあるものがあります。メチオニンは例外で、一文字目がA、二文字目がTで三文字目がGである、ただ一個の“ATGコドン”だけがこれに対応しています。全てのタンパクがメチオニンから始まりますので、これがいくつもあると混乱の元になるからだろうと思われます。このほかにトリプトファンのコドンも一つだけです。それ以外に対応するアミノ酸がなく、そこでタンパクの合成が止まってしまう“停止コドン”が3個あります。どこまでも遺伝子を読み続けると、とてつもなく長いアミノ鎖ができてしまいますので、決まった場所に停止コドンがあることが必要なのです。
このほかにも、考えておかなければいけないことがあります。3個の塩基の順列が20個のアミノ酸を規定していて、これを“コドン”といっています。一文字目、二文字目と三文字目にそれぞれT、C、AとGの4通りがありますから、4を3回掛けた回数、すなわち64個の組み合わせがあります(図4)。アミノ酸の総数は20個しかありませんから、殆どのアミノ酸に複数のコドンが対応し、中には同じアミノ酸の合成を指令するコドンが6種類もあるものがあります。メチオニンは例外で、一文字目がA、二文字目がTで三文字目がGである、ただ一個の“ATGコドン”だけがこれに対応しています。全てのタンパクがメチオニンから始まりますので、これがいくつもあると混乱の元になるからだろうと思われます。このほかにトリプトファンのコドンも一つだけです。それ以外に対応するアミノ酸がなく、そこでタンパクの合成が止まってしまう“停止コドン”が3個あります。どこまでも遺伝子を読み続けると、とてつもなく長いアミノ鎖ができてしまいますので、決まった場所に停止コドンがあることが必要なのです。
もうひとつ気がつくことがありませんか? とても大切なことです。それぞれのアミノ酸が、一文字目と二文字目の塩基で決まってしまい、三文字目の塩基はTとC(いずれもピリミジンです)、あるいはAとG(両方ともプリンです)のどちらであっても、同じアミノ酸ができてしまうことです。TからC(CからTでも)、あるいはAからG(GからAでも)に置き替わっても同じアミノ酸ができるこのような塩基の置換を「同義置換」といっています。二文字目は、どのような置換でもアミノ酸が違ってきますので、すべてが「異義置換」となります。同様に、一文字目の置換も、ロイシンとアルギニンの例外をのぞいて、ほか全部が異義置換となります。
ある場所の塩基が入れ替わったときに、それによってアミノ酸が変わってしまいますと、合成されるタンパクに影響がでて、構造と機能が失われることが多いのです。ですから、異義置換を起こす変異があるとそのウイルスは生きていけないことになります。すなわち異義置換はこのように淘汰の影響を受けるので、ウイルスの固有の変異率を正確に反映できないことになります。したがって、たとえ変異してもアミノ酸が変わらない塩基の「同義置換」だけが、ウイルスの進化率を計算する基準になります。B型肝炎ウイルスには約3,200個の塩基がありますが、遺伝子を1個だけ担っている部分は約半分(1,600個)しかなく、しかも三文字目の核酸はその3分の1の約500個あまりしか存在しないことになります。すなわち変異率を計算できる塩基の数は、わずかに全配列の約6分の1しかないことになります。
分子時計から計算した宿主とB型肝炎ウイルスの起源
同一人に感染していたB型肝炎ウイルスの「同義置換」部位に相当する箇所で、ある期間(たとえば10年間)に塩基が何個入れ替わったかを調べることによって、年間の変異率を計算できます。しかし、たとえ「同義置換」が起こっても、長年の間にそれがもう一度「同義置換」すれば元の塩基に戻ってしまいますので、見かけ上は変異が起こらなかったことになります。そのことも考慮に入れて、B型肝炎ウイルスの年間・変異率は約10万分の1(10のマイナス5乗)と計算されています。この変異率を頼りにすれば、二個のB型肝炎ウイルス間で違っていた該当する塩基数をそれで割ることによって、何年前にその二つのウイルスが別の進路をとるようになったのか、すなわちいつ分化したかを、計算することができます。
 宿主である動物の遺伝子(DNA配列)変異率はもっともっと低く10億分の1(10のマイナス9乗)ほどです。B型肝炎ウイルスは、ヒトだけでなくチンパンジーに代表される野生の類人猿にも感染しています。人類とチンパンジーは同じ祖先から分化して、とても近い関係にありますのでヒトB型肝炎ウイルスはチンパンジーに感染しますし、また逆にチンパンジーのB型肝炎ウイルスもヒトに感染するだろうと考えられています。これ以外にも、野生のリス(ウッドチャックなど)とトリ(アヒル、サギ、ツルなど)にも、その動物固有のB型肝炎ウイルスが持続的に感染していることが知られています。
宿主である動物の遺伝子(DNA配列)変異率はもっともっと低く10億分の1(10のマイナス9乗)ほどです。B型肝炎ウイルスは、ヒトだけでなくチンパンジーに代表される野生の類人猿にも感染しています。人類とチンパンジーは同じ祖先から分化して、とても近い関係にありますのでヒトB型肝炎ウイルスはチンパンジーに感染しますし、また逆にチンパンジーのB型肝炎ウイルスもヒトに感染するだろうと考えられています。これ以外にも、野生のリス(ウッドチャックなど)とトリ(アヒル、サギ、ツルなど)にも、その動物固有のB型肝炎ウイルスが持続的に感染していることが知られています。
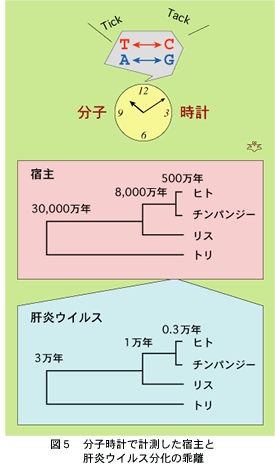
宿主である動物で遺伝子を構成する塩基一個あたりの変異率がわかっていれば、他種動物で対応する遺伝子配列の塩基が何個違うかを調べ、その数を変異率で割ることによって、それぞれの動物が何年昔に分化したかを推定することができます。この手法を「分子時計」といっています(図5)。これによってヒトとチンパンジーがそれぞれ別の道をたどり始めたのは5百万年前であると推定されています。また、ヒトがリス等の齧歯類から分化したのは8千万年前、トリと分かれたのは3億年以前(!)という気の遠くなるような昔の出来事と推定されています。
一方おなじ「分子時計」を各種動物のB型肝炎ウイルスに当てはめますと、ヒトのB型肝炎ウイルスとチンパンジーのB型肝炎ウイルスとが「分化」したのは、たったの3千年以前だったという結果になります。同様にヒトとリスのB型肝炎ウイルスは1万年前、ヒトとトリのB型肝炎ウイルスは3万年以前に分化してそれぞれ別の進路をとった、という計算にもなります。宿主であるそれぞれの動物での分化時期と比べますと、千分の一から万分の一ほどもの大きな違いがあり、B型肝炎ウイルスは宿主よりずっと最近になってから「分化」した、という結論になります。
今この結果をどのように解釈するかを巡って、B型肝炎ウイルス研究をしている専門家の間でも、意見が大きく分かれているのです。
人類発祥の地アフリカから五大陸へ向かった人類移動の歴史
B型肝炎ウイルスの起源を知るためには、人類がどのように発祥して旧世界から新世界にいつ渡来したかを知る必要があります。併せて、他動物とりわけ類人猿のB型肝炎ウイルスに関する情報を分析し、その結果からヒトと類人猿(特にチンパンジー)との間で、過去のどの時点で種を越えた「交差感染」によるB型肝炎ウイルスのやりとりがあったのかをも、推定しなければなりません。
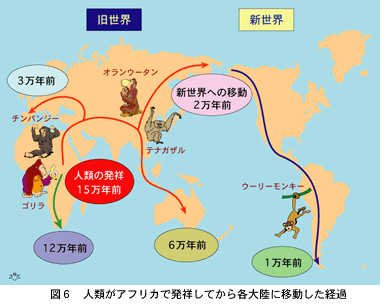 現世の人類は、約15万年以前にアフリカの中部で進化したと考えられています(図6)。そこから南北の二方向に移動して、アフリカ南端には12万年以前に到着しましたが、北方への道は全世界に通じていたのです。ユーラシア大陸に到着してからは、アジアとヨーロッパの東西二方向に分かれ、アジアからは南にオーストラリアまで、北にはベーリング海峡に達しました。そして、ベーリング海峡が陸続きであった2万年以前には新大陸に渡り、北アメリカを経て南アメリカの先端に到達したのは1万年前のことです。これは、勿論有史前の先住民がたどった足跡で、戦略と貿易を目的とした有史後の民族移動は含みません。
現世の人類は、約15万年以前にアフリカの中部で進化したと考えられています(図6)。そこから南北の二方向に移動して、アフリカ南端には12万年以前に到着しましたが、北方への道は全世界に通じていたのです。ユーラシア大陸に到着してからは、アジアとヨーロッパの東西二方向に分かれ、アジアからは南にオーストラリアまで、北にはベーリング海峡に達しました。そして、ベーリング海峡が陸続きであった2万年以前には新大陸に渡り、北アメリカを経て南アメリカの先端に到達したのは1万年前のことです。これは、勿論有史前の先住民がたどった足跡で、戦略と貿易を目的とした有史後の民族移動は含みません。
人類が各大陸に移動するずっと以前に、野生動物が同じ経路で各地に拡散しただろうと想像できます。特に類人猿の移動と進化が、B型肝炎ウイルスの人類感染にとって最も重要であろうと考えられます。人類が農耕を始めたのは、僅かに5千年以前のことで、それまでは狩猟が主な生存手段だったのでしょう。特に火を入手した1万年以前からは、食肉調理の方法が大幅に改良したはずです。
地域に依存したB型肝炎ウイルスの遺伝子型
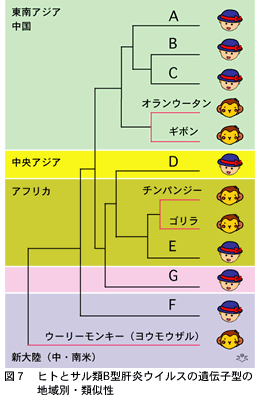 ヒトB型肝炎ウイルスは、3,200個の塩基配列が8%(約260個)以上違うことによって、アルファベット大文字のAからGであらわされる7種類の遺伝子型(ゲノタイプ)に分類されています(第六話をご覧下さい)。B型肝炎ウイルスは旧大陸の大型類人猿である野生のチンパンジーとゴリラにも持続感染していて、それぞれチンパンジーB型肝炎ウイルスおよびゴリラB型肝炎ウイルスと名付けられています。東南アジア地方に棲息しているオランウータンもB型肝炎ウイルスに感染していますし、もっと広範囲に分布するテナガザル(ギボン)もギボンB型肝炎ウイルスに感染しています。これら類人猿のB型肝炎ウイルスは、ヒトB型肝炎ウイルスにとてもよく配列が似ています。それどころか7種類あるヒトB型肝炎ウイルス間の塩基配列の違い(8%)ほども違っていない霊長類B型肝炎ウイルスが存在します(図7)。系統発生学的にヒトB型肝炎ウイルスと見分けがつかないので、同一ウイルスと考えるほかなさそうです。
ヒトB型肝炎ウイルスは、3,200個の塩基配列が8%(約260個)以上違うことによって、アルファベット大文字のAからGであらわされる7種類の遺伝子型(ゲノタイプ)に分類されています(第六話をご覧下さい)。B型肝炎ウイルスは旧大陸の大型類人猿である野生のチンパンジーとゴリラにも持続感染していて、それぞれチンパンジーB型肝炎ウイルスおよびゴリラB型肝炎ウイルスと名付けられています。東南アジア地方に棲息しているオランウータンもB型肝炎ウイルスに感染していますし、もっと広範囲に分布するテナガザル(ギボン)もギボンB型肝炎ウイルスに感染しています。これら類人猿のB型肝炎ウイルスは、ヒトB型肝炎ウイルスにとてもよく配列が似ています。それどころか7種類あるヒトB型肝炎ウイルス間の塩基配列の違い(8%)ほども違っていない霊長類B型肝炎ウイルスが存在します(図7)。系統発生学的にヒトB型肝炎ウイルスと見分けがつかないので、同一ウイルスと考えるほかなさそうです。
AからFまでのヒトB型肝炎ウイルス遺伝子型の分布は地域によって違っていて(G型遺伝子の報告はまだ少なく、世界分布がわかっていません)、アフリカでは遺伝子型Eが多いのです。そして、同じくアフリカに有史以前から住んでいるチンパンジーとゴリラのB型肝炎ウイルスと、遺伝子型EのヒトB型肝炎ウイルスとはとても近い関係にあります。同じような現象が、東南アジアと中国に多い(日本にもですが)遺伝子型BとCのヒトB型肝炎ウイルスと土着しているオランウータンとギボンのB型肝炎ウイルスとの間にも見られます。それほど緊密な関係ではないのですが、新大陸に棲息している下等なサル類であるウーリーモンキー(ヨウモウザル)に感染しているウーリーモンキーB型肝炎ウイルスは、同じく中南米にだけ分布している遺伝子型FのヒトB型肝炎ウイルスにかなり近い関係にあります。
このようにAからFまでのヒトB型肝炎ウイルス遺伝子型は、おそらく同じ場所で人類にさきがけて長期間棲息していたであろう類人猿B型肝炎ウイルスと緊密な関係があります。どの時点とは断言できませんが、人類が発祥した古代のアフリカでも、また人類が各大陸に渡来してからも、狩猟と動物の解体および食肉を通じて先住の類人猿から彼ら固有のB型肝炎ウイルスがまずヒトに感染して、それ以後二次的にヒトの間に広まった可能性があります。
チンパンジー亜種に感染しているB型肝炎ウイルスの遺伝子亜型
「分子時計」の結果を額面通り受け取るのであれば(図5)、ヒトのB型肝炎ウイルスはチンパンジーB型肝炎ウイルスと時を同じくして、僅か3千年前に分化し始めて、現在確認されているAからGまでの7種類の遺伝子型に分かれた、という結論となります。また、下等の猿類であるウーリーモンキーのウイルスに一番近い関係にある遺伝子型FのB型肝炎ウイルスが系統発生学上もっとも原始的なので、これを根拠として新大陸がB型肝炎ウイルス発祥の地であろう、と推論できないこともありません。
事実、そのような路線に基づいて、B型肝炎ウイルスは3千年以前に南アメリカに発生し、その後の貿易と奴隷輸入によって他の四大陸に広まった、と主張しているウイルス学者もいます。でも、本当にそう考えてよいのでしょうか?
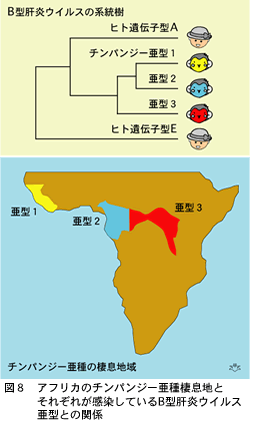 最近、この問題を解く鍵が見つかりました。チンパンジーはアフリカの赤道直下にある帯状の地域に棲息していますが、三つの亜種があります(図8)。チンパンジーの生態を根気よく観察した結果、驚くべき事実が明らかになりました。亜種は150万年以前に分化して、それ以来お互いに混合することなく、それぞれが純粋に保たれている、ということが判明したのです。住みかに遺された体毛を採取してそれから“ミトコンドリアDNA”を抽出後、増幅して系統発生学的に研究する技術が進んだので、チンパンジーの遺伝学的研究の素材が、大幅に増加しました。捕まえて採血をする必要がなくなりチンパンジーに多大な迷惑をかけないでもすむようになったのです。また三つのチンパンジー亜型は、遺伝学的に大きく違っていますので、それぞれが独立した種であると考えた方が良いくらいなのです。
最近、この問題を解く鍵が見つかりました。チンパンジーはアフリカの赤道直下にある帯状の地域に棲息していますが、三つの亜種があります(図8)。チンパンジーの生態を根気よく観察した結果、驚くべき事実が明らかになりました。亜種は150万年以前に分化して、それ以来お互いに混合することなく、それぞれが純粋に保たれている、ということが判明したのです。住みかに遺された体毛を採取してそれから“ミトコンドリアDNA”を抽出後、増幅して系統発生学的に研究する技術が進んだので、チンパンジーの遺伝学的研究の素材が、大幅に増加しました。捕まえて採血をする必要がなくなりチンパンジーに多大な迷惑をかけないでもすむようになったのです。また三つのチンパンジー亜型は、遺伝学的に大きく違っていますので、それぞれが独立した種であると考えた方が良いくらいなのです。
野生のチンパンジーにもB型肝炎ウイルスが感染しています(図8)。それぞれの亜種に感染しているB型肝炎ウイルスはすこしずつ違っていても、その塩基配列差はウイルス全長で8%に達しませんから遺伝子型は同じだと云うことになります。ヒトのAからGまでの遺伝子型間の違いほどの差は見られないのです。
この結果をどのように受け止めたらよいのでしょうか? 人類の進化はアフリカ中央で15万年以前まで続いていました。チンパンジーも勿論その時代に同じ場所に棲息していました。そしておそらくB型肝炎ウイルスの祖先もそのころから存在しいて人類とチンパンジーに人畜共通感染していた可能性があります。チンパンジー固有のB型肝炎ウイルスは、ヒトB型肝炎ウイルスの中でアフリカ原住民に広く見られる遺伝子型E・ウイルスに最も近縁にあることからも、こう考えて良いでしょう。
以来、人類はアフリカから広く四大陸に移動し動物界を制覇しました。その先々で野生の類人猿を狩猟し摂食する課程で彼らに感染していた土地固有のB型肝炎ウイルスに感染する機会があったものと思われます。その総合的な結果として、人類の中で7種の遺伝子型を持ったB型肝炎ウイルスが進化しました。
一方チンパンジーの方は、その間ずっとアフリカの狭い範囲にとどまっていました。そして、三カ所の違った場所で150万年もかけて異なる亜種に進化したのです。同じ大陸の同じ緯度(赤道直下)で、三つの亜種が独立して生存し、雑種ができなかったのは不思議な話です。これはチンパンジーの特殊な生態と社会性のためだろう、と想像されています。チンパンジーは泳ぐことができません。たとえば川ができれば永遠に隔離されてしまいますし、そのような出来事がアフリカの歴史にもあったのではないでしょうか。しかしその間B型肝炎ウイルスは殆ど進化せず、いま「生きた化石」として真実を語りかけているのです。
B型肝炎ウイルス:ヒトと他動物間で「ウイルス・やりとり」の軌跡
人類の健康を脅かしている多くの病原性ウイルス感染が、他動物からうつった「人畜共通感染」の結果であることが、最近注目されています。典型例はHIV(human
immunodeficiency virus)感染で、約30年以前から少なくとも7回にわたってアフリカのチンパンジーから、血液を介して現地の食肉解体業者に感染し、それが瞬く間に全世界に拡散したと考えられています。HIVに対応した猿のウイルス(simian
immunodeficiency virus [SIV])が全世界にひろまっていて、調べられただけで30種類もの野生類人猿に感染しています。ウイルスは自然の宿主では殆ど害をしませんが、宿主が変わるとHIV感染のように重い病気を起こすことがあります。B型肝炎ウイルスも、チンパンジーを初めとしたサル類ではヒトのように重い肝臓病は起こしませんので、おそらく彼らが自然宿主であろうと思われます。
肝炎ウイルスの中でも、E型肝炎ウイルスは広く養豚に蔓延し、野生のイノシシとシカにも感染しています。動物からヒトへ、またはその逆方向の感染経路がありそうです。C型肝炎ウイルスの歴史は浅く、人類に感染してからまだ200年位しかたっていないと考えられています。B型肝炎ウイルスと違って、他動物には対応するC型肝炎ウイルスはまだ発見されていませんが、おそらく過去のある時点で人畜共通感染によって動物から人類にうつったのだろう、と想像できます。二十一世紀に入ってからも既にいくつかの「新興感染症」が発生して、その殆どで「人畜共通感染症」の経路が疑われています。SARS(severe
acute respiratory syndrome)ウイルスとトリ・インフルエンザウイルスなど、記憶に新しいところです。
HIVのサル版であるSIVほどよく調べられていませんが、B型肝炎ウイルスも広く全世界の野生動物、特にサル類に感染している可能性があり今後詳しく調べる必要があります。その研究で得られた結果の系統発生学的解析からB型肝炎ウイルスが人類全般にわたって、いつどこでどのように拡散したかが次第に明らかになることを期待できます。
![]()